Chlorarachniophyte algae [2] ( lat. Chlorarachnea ) is a treasure of marine unicellular algae , which is part of the group of tserkozoey , which in recent studies systematists assign the rank of class and type, respectively [3] . Distributed in the seas of the tropical and temperate zones. The organisms of this group are characterized by the presence of four- membrane chloroplasts , in the periplastid space of which there is a nucleomorph — the reduced nucleus of endosymbiotic eukaryote [2] . Chlorarachniophyte algae are mixotrophs ; they contain chlorophylls a and b and are capable of phagotrophic nutrition. Propagate vegetatively and asexually, some have described the sexual process . According to the latest data, the treasure contains 14 species in 8 genera.
| Chlorarachniophyte algae | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
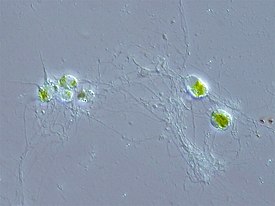 Chlorarachnion reptans | |||||||||||
| Scientific classification | |||||||||||
| |||||||||||
| International scientific name | |||||||||||
Chlorarachnea Cavalier-Smith, 1993 | |||||||||||
| Synonyms | |||||||||||
| |||||||||||
| Subsidiary taxa | |||||||||||
see text | |||||||||||
In the botanical taxonomy of the clade, the class Chlorarachniophyceae corresponds. The obsolete name - chlorarachniophytes ( lat. Chlorarachniophyta ), given even at the first description of the clade as a department of algae [4] [2] , is also widespread in the literature.
Content
- 1 Study History
- 2 Cell Biology
- 3 Reproduction and the life cycle
- 4 Distribution and ecology
- 5 Phylogeny and classification
- 6 notes
- 7 Literature
Study History
The first known species of chlorarachniophyte, Chlorarachnion reptans , has been described from a culture of siphon green algae harvested in the Canary Islands . Initially, it was attributed to the yellow-green algae in the composition of Stramenopila . In 1966, Chlorarachnion reptans was rediscovered in Mexico and isolated in pure culture. The department of Chlorarachniophyta was isolated only in 1984 on the basis of the ultrastructure and pigment composition of Chlorarachnion reptans [4] [5] . The class name is given according to the type genus Chlorarachnion (from other Greek: χλωρός “green” and ἀράχνη “web”) [2] .
Cell Biology
Chlorarachniophyte cells are flagellates or amoeboid with filamentous pseudopodia . Cell size varies from 5 to 100 microns . Bigellowiella longifila can form floating mucous colonies [6] . The cells are naked, mononuclear, but there are trichocysts . The flagellum stages have one smooth flagellum wrapped in a spiral around the cell. Chlorarachnion is characterized by the development of reticulopodia (filopodia), which merge in neighboring cells with the formation of reticular plasmodium (meroplasmodium, or reticular plasmodium) [7] . Up to hundreds of cells can be combined in the composition of meroplasmodium. In other chloroarachniophytes, meroplasmodium does not develop, only in Cryptochlora perforans the filopodia of two or three cells can merge [2] .
Crista mitochondria are tubular [8] . Sequenced mitochondrial DNA chlorarachniophyte is characterized by some features. So, the codons GUG and UUG can act as initiators , along with AUG. There is evidence of gene transfer from the mitochondrial genome to the host nuclear genome in the process of species divergence. Moreover, the preservation of the order of genes ( synteny ) is rather low, which indicates frequent rearrangements in the mitochondrial genomes [9] .
Chloroplasts chloroarachniophyte contain chlorophylls a and b , the nature of carotenoid pigments is unknown. Chloroplasts are surrounded by four membranes , of which two are proprietary , and the other two belong to the chloropast endoplasmic reticulum, and the second and third membranes can merge for some extent. It is believed that the second membrane is the plasmalemma of the endosymbiont, and the fourth membrane is derived from the host. The outer chloroplast membrane does not bind to the outer nuclear membrane and does not carry ribosomes . Lamellas consist of 1-3 thylakoids , there is no encircling lamella. Pyrenoids are found in plastids of all chloroarachniophytes, except for Partenskyella glossopodia [10] . Each pyrenoid is clearly directed toward the center of the cell; by this sign, chloroarachniophyte can be easily distinguished from other chlorophyll b-containing algae when viewed under a light microscope . The prominent end of the pyrenoid always has a capping vesicle , in which a reserve product is accumulated - β-1-3-glucan (possibly ) [6] [11] .
Plastid genomes of chloroarachniophyte, sequenced to date, are conservative in size, number of genes, and gene order. Apparently, the plastid genome of the endosymbiont, swallowed by chloroarachniophytes, practically did not change during the divergence of species [12] .
The mechanism of protein delivery to plastid in chlorarachniophyte is significantly different from other plastids resulting from secondary endosymbiosis. As in other algae with plastids, which are the result of secondary endosymbiosis, in chloroarachniophyte, plastid proteins have a two-part N-terminal signal of plastid localization, consisting of special SP and TPL sequences. This testifies to the presence of special translaconones in chloroarachniophyte that ensure the passage of proteins of two internal membranes, but it turned out that these complexes are significantly different from those of other algae with secondary endosymbiosis. Moreover, it was shown that the mechanism of protein transport through the second outer membrane is fundamentally different [13] .
Of particular interest in the structure of cells of chloroarachniophyte algae is nucleomorph, the reduced nucleus of the endosymbiont, which gave rise to chloroplast. In chloroarachniophyte, green algae became the endosymbiont (according to the composition of xanthophylls - from the number of , according to molecular phylogenetics and ecology - ulvofitsievye and trebuksiofitsievyh ). The nucleomorph is located in the periplastid space (between the second and third membranes), in which the endosymbiont cytoplasm and eukaryotic ribosomes are also present. It is surrounded by two membranes with pores, contains DNA and a structure close to the nucleolus , eukaryotic type ribosomes and 3 small linear A T- enriched chromosomes . The length of the nucleomorph chromosomes varies from 90 to approximately 210 kilobases (kb), and the total size of the nucleomorph genome varies from 330 kb in Lotharella amoebiformis to 610 kb in the unidentified strain of CCMP622 [14] . The density of genes in nucleomorphs is very high (sometimes genes even overlap), although they contain a certain number of very short introns . RRNA gene series are located in regions. Proteins encoded by a nucleomorph serve it itself, only a few function in chloroplasts. In Bigelowiella natans, histones and are encoded by the nucleomorph itself, and the genes of histones and transferred to the host nucleus. Linker histones ( ) are completely absent in nucleomorphs. There are some differences in the post-translational modifications of the histones ( ) of the nucleomorph and nucleus. Thus, heterochromatin labels in the form of H3K9 and H3K27 methylation, H3K36 methylation corresponding to transcription elongation, almost all transcriptional activation / repression labels, and almost completely no phosphorylation associated with the dynamics of chromosomes 15 mit ] .
The nucleomorph divides before dividing the chloroplast by invading the membranes of its membrane. No mitotic spindle was found there; therefore, the mechanism of chromosome divergence between daughter nucleomorphs is not clear [11] . It was shown that FtsZD proteins are partially involved in the formation of constriction near the inner pair of membranes, however, the mechanism of constriction formation in the outer part of the membranes is unknown [16] .
In addition to chlorarachniophyte, cryptophytic algae have the nucleomorph with three chromosomes of the same organization, but they have a different origin from red algae. Thus, the presence of nucleomorphs in representatives of these groups can be considered the result of convergent evolution [17] . So, there are four genomes in the cells of chlorarachniophyte and cryptophyte: the nuclear, plastid, mitochondrial and the genomorphs. Both the chlorarachniophyte alga Bigelowiella natans and the cryptophyte alga were shown to have the haploid nuclear genome, the diploid nucleomorphic gene in B. natans and the tetraploid genome in G. theta , and the plastid and mitochondrial genomes in both species have both each corresponding organelle [18] .
Reproduction and Life Cycle
Chlorarachniophyte is characterized by vegetative propagation by cell division in two, asexual using zoospores and sexual ( iso - and anisogamy ). In the life cycle , amoeboid, coccoid and monad stages can occur. All three stages are present only in Chlorarachnion reptans and Lotharella amoeboformis . Coccoid stages have a multilayer cell wall and are often considered as cysts [17] . In Chlorarachnion reptans , which has meroplasmodium, in conditions of lack of nutrients, stellate vegetative cells draw in reticulopodia, round off and dress on a dense cell wall. Resting cells exist only due to photosynthesis and are not capable of heterotrophy . Under favorable conditions, resting cells again turn into stellate vegetative cells with reticulopodia. The formation of zoospores occurs by twofold division of a resting cell. As a result, single-flagged zoospores are formed, the flagellum of which is twisted around the back of the floating cell. Flagellum carries thin lateral hairs and departs from the secondarily unpaired single kinetosome . During sexual reproduction, a stationary female gamete merges with a movable stellate male gamete. The zygote formed during fertilization is divided by meiosis and sprouts by a stellate vegetative cell [19] [20] . Lotharella globosa showed significant morphological diversity (the presence or absence of certain stages in the life cycle) within the species between different cultured strains [21] .
The behavior of daughter cells formed during vegetative propagation may vary. For example, in Lotharella vacuolata, a coccoid cell undergoes division, one of the daughter cells takes an amoeboid shape and floats away, and the other daughter cell remains inside the mother. In the planktonic alga Bigellowiella longifila, the flagellate and amoeboid stages can be vegetative stages, and the amoeboid stage has only one long filopodia. When an amoeboid cell divides, one of the daughter cells inherits filopodia and swims away, pumping the contents of the cell to the far end of the filopodia, while the other remains stationary and forms filopodia only after some time [22] .
Distribution and Ecology
Being the only group of algae that have never been found in fresh water [23] , chloroarachniophyte species live in the warm seas of the tropical and temperate zones around the world. Amoeboid and coccoid forms live in coastal waters, and monadic forms are represented in oceanic picoplankton . Chlorarachniophytes were found in a wide variety of marine habitats: on sandy beaches, rocky shores, in the tidal zone, near the surface of the water, on the surface of marine plants , on grains of sand and near the bottom [23] . Chlorarachniophytes are mixotrophs ; they are capable of photosynthesis, but can also feed on bacteria , flagellates, and eukaryotic algae [24] .
Phylogeny and classification
Initially, the clade was described as a department of the algae Chlorarachniophyta, consisting of one genus Chlorarachnion [4] , and subsequently the number of genera increased. Molecular analysis confirmed the monophilia of the taxon. Analysis of rRNA genes , tubulin and actin subunits showed that chlorarachniophytes are closest to heterotrophic amoeboflagellates — and , together with which (as well as plasmodiophore ) form the Cercozoa group [25] . However, the group closest to the host cells of chloroarachniophyte could not be determined. Cavalir-Smith suggested that chlorarachniophytes and euglenophytes come from a single photosynthetic ancestor, but further study has shown that this assumption is incorrect [26] [27] . After the approval of the tserkozoys in the type rank (before that, some scholars raised the rank of the group to the kingdom inclusively [2] ), the rank of chloroarachniophyte algae was reduced to class, synonymous with the names of the department and class. Since tserkozoey taxa are described mainly by the ICZN rules, the older treasure taxa were renamed in accordance with the rules of this code [3] [1] .
The differences between the genera of the class lie in the structural features of the pyrenoid and the location of the nucleomorph, however, the exact relationships between the individual genera are unknown [25] . As of June 2016, 8 genera and 14 species are included in the class of chloroarachniophyte algae [21] [28] [29] :
- Class Chlorarachnea [ Chlorarachniophyceae Hibberd & Norris, 1984 , orth. var.]
- Order Chlorachniida [ Chlorarachniales Ishida & Y. Hara, 1996 , orth. var.]
- Chlorarachniaceae Family
- Amorphochlora A. Yabuki & S. Ota, 2011 (1 view)
- Bigelowiella Moestrup, 2001 (2 species)
- Chlorarachnion Geitler, 1930 (1 view)
- Cryptochlora E. Calderon-Saenz & R. Schnetter, 1987 (1 view)
- Gymnochlora K. Ishida & Y. Hara, 1996 (2 species)
- Lotharella K. Ishida & Y. Hara, 1996 (5 species)
- Norrisiella S. Ota, K. Ueda & K. Ishida, 2007 (1 species)
- Partenskyella S. Ota, D.Vaulot, FLGall, A.Yabuki & K.Ishida, 2009 (1 species)
- Chlorarachniaceae Family
- Order Chlorachniida [ Chlorarachniales Ishida & Y. Hara, 1996 , orth. var.]
Notes
- ↑ 1 2 3 Class Chlorarachnea (English) in the World Register of Marine Species . (Retrieved July 19, 2016) .
- ↑ 1 2 3 4 5 6 Belyakova et al., 2006 , p. four.
- ↑ 1 2 Ruggiero MA , Gordon DP , Orrell TM , Bailly N. , Bourgoin T. , Brusca RC , Cavalier-Smith T. , Guiry MD , Kirk PM A Higher Level Classification of All Living Organisms // PLoS ONE . - 2015. - Vol. 10, no. 4. - P. e0119248. - DOI : 10.1371 / journal.pone.0119248 . - PMID 25923521 .
- ↑ 1 2 3 David J. Hibberd, Richard E. Norris. Cytology and ultrastructure of Chlorarachnion reptans (Chlorarchniophyta Divisio nova, Chlorachniophyceae Classis nova). (English) // Journal of Phycology. - 1984. - Vol. 20, no. 2 . - P. 310-330.
- ↑ Brodie, Lewis, 2007 , p. 178.
- ↑ 1 2 Brodie, Lewis, 2007 , p. 173.
- ↑ Houseman et al., 2010 , p. 180.
- ↑ Belyakova et al., 2006 , p. 5.
- ↑ Tanifuji G. , Archibald JM , Hashimoto T. Comparative genomics of mitochondria in chlorarachniophyte algae: endosymbiotic gene transfer and organellar genome dynamics. (English) // Scientific reports. - 2016. - Vol. 6. - P. 21016. - DOI : 10.1038 / srep21016 . - PMID 26888293 .
- ↑ Ota S. , Vaulot D. , Le Gall F. , Yabuki A. , Ishida K. Partenskyella glossopodia gen. et sp. nov., the first report of a Chlorarachniophyte that lacks a pyrenoid. (English) // Protist. - 2009. - Vol. 160, no. 1 . - P. 137-150. - DOI : 10.1016 / j.protis.2008.09.003 . - PMID 19013103 .
- ↑ 1 2 Belyakova et al., 2006 , p. 5-7.
- ↑ Suzuki S. , Hirakawa Y. , Kofuji R. , Sugita M. , Ishida K. Plastid genome sequences of Gymnochlora stellata, Lotharella vacuolata, and Partenskyella glossopodia reveal remarkable structural conservation among chlorarachniophyte species. (англ.) // Journal of plant research. — 2016. — Vol. 129, no. 4 . — P. 581—590. — DOI : 10.1007/s10265-016-0804-5 . — PMID 26920842 .
- ↑ Hirakawa Y. , Burki F. , Keeling PJ Genome-based reconstruction of the protein import machinery in the secondary plastid of a chlorarachniophyte alga. (англ.) // Eukaryotic cell. — 2012. — Vol. 11, no. 3 . — P. 324—333. — DOI : 10.1128/EC.05264-11 . — PMID 22267775 .
- ↑ Silver TD , Koike S. , Yabuki A. , Kofuji R. , Archibald JM , Ishida K. Phylogeny and nucleomorph karyotype diversity of chlorarachniophyte algae. (англ.) // The Journal of eukaryotic microbiology. - 2007. - Vol. 54, no. 5 . — P. 403—410. — DOI : 10.1111/j.1550-7408.2007.00279.x . — PMID 17910684 .
- ↑ Marinov GK , Lynch M. Conservation and divergence of the histone code in nucleomorphs. (англ.) // Biology direct. — 2016. — Vol. 11, no. 1 . — P. 18. — DOI : 10.1186/s13062-016-0119-4 . — PMID 27048461 .
- ↑ Hirakawa Y. , Ishida K. Prospective function of FtsZ proteins in the secondary plastid of chlorarachniophyte algae. (англ.) // BMC plant biology. — 2015. — Vol. 15. — P. 276. — DOI : 10.1186/s12870-015-0662-7 . — PMID 26556725 .
- ↑ 1 2 Белякова и др., 2006 , с. 7.
- ↑ Hirakawa Y., Ishida K. Polyploidy of endosymbiotically derived genomes in complex algae // Genome Biology and Evolution. — 2014. — Vol. 6, no. 4. — P. 974—980. — DOI : 10.1093/gbe/evu071 . — PMID 24709562 .
- ↑ Lee, 2008 , p. 319.
- ↑ Хаусман и др., 2010 , с. 180—181.
- ↑ 1 2 Hirakawa Y. , Howe A. , James ER , Keeling PJ Morphological diversity between culture strains of a chlorarachniophyte, Lotharella globosa. (English) // Public Library of Science ONE. — 2011. — Vol. 6, no. 8 . — P. e23193. — DOI : 10.1371/journal.pone.0023193 . — PMID 21858028 .
- ↑ Brodie, Lewis, 2007 , p. 177—178.
- ↑ 1 2 Brodie, Lewis, 2007 , p. 172.
- ↑ Белякова и др., 2006 , с. 7—8.
- ↑ 1 2 Белякова и др., 2006 , с. 6, 8.
- ↑ Tolweb: Chlorarachniophytes .
- ↑ Rogers MB , Gilson PR , Su V. , McFadden GI , Keeling PJ The complete chloroplast genome of the chlorarachniophyte Bigelowiella natans: evidence for independent origins of chlorarachniophyte and euglenid secondary endosymbionts. (англ.) // Molecular biology and evolution. - 2007. - Vol. 24, no. 1 . — P. 54—62. — DOI : 10.1093/molbev/msl129 . — PMID 16990439 .
- ↑ Семейство Chlorarachniaceae (англ.) в Мировом реестре морских видов ( World Register of Marine Species ). (Проверено 19 июля 2016) .
- ↑ Family: Chlorarachniaceae (англ.) . AlgaeBase . (Проверено 19 июля 2016) .
Literature
- Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. Ботаника: в 4 т. Т. 2. — М. : Издат. центр «Академия», 2006. — 320 с. — ISBN 978-5-7695-2750-1 .
- Хаусман К., Хюльсман Н., Радек Р. Протистология / Пер. from English С. А. Карпова. Ed. С. А. Корсуна. — М. : Товарищество научных изданий КМК, 2010. — 495 с. — ISBN 978-5-87317-662-5 .
- Lee R. E. Phycology. 4th ed . — New York: Cambridge University Press, 2008. — 561 p. — ISBN 978-0-521-68277-0 .
- Juliet Brodie, Jane Lewis. Unravelling the algae: the past, present, and future of algal systematics . — London, UK: CRC Press, 2007. — 420 p. — ISBN 978-0-8493-7989-5 .