Endospores are resting forms of bacteria that are formed as a result of the coordinated differentiation of two daughter cells formed by doubling the genome of the original bacterium, and one of them penetrates the cytoplasm of the other daughter cell, which becomes external. Further, the external daughter cell (sometimes called sporangia ) dies by programmed death , and the internal cell (prespore) becomes the endospore itself and enters a state of maximum physiological rest, in which all physiological processes inside the spore stop ( hyperanabiosis ). It becomes extremely resistant to adverse environmental conditions and can remain viable for a long time [1] .
Two cases are described when the mother cell gives not one but two endospores. Thus, the large (35 μm long) uncultured bacterium Metabacterium polyspora , which lives in the gastrointestinal tract of some rodents , forms up to nine endospores per sporangia. Another giant uncultured bacterium, Epulopiscium sp. (it can reach a size of 60 × 300 μm), which lives inside a tropical unicorn fish , forms two endospores in one sporangia [2] .
Content
Sporogenous bacteria
Endospores form representatives of about 25 genera of the firmicut type: Bacillus , Clostridium , , , , , Heliophilum , Oscillospira , Sporobacter , , , , Sporotomaculum , , Thermoactinomyces and others. In the vast majority of cases, sporogenous bacteria are gram-positive , although there are exceptions. For example, and Heliophilum fasciatum are among the gram-negative bacteria [3] .
Endospore structure
The main structural elements of the mature endospore:
- cytoplasmic compartment, or core;
- combined cell wall consisting of an inner wall and an outer wall (cortex);
- endospore cytoplasmic membrane (inner membrane );
- cytoplasmic membrane sporangia (outer membrane);
- internal and external spore integument;
- outer cover ( exosporium ) [4] .
The location of endospores within the mother cell may be different, and the type of spore formation in some cases is used to identify an unknown culture. Sometimes the forming endospore reaches such sizes that it expands the sporangium in the middle or at one of the ends [5] .
The endospore core contains a chromosome and small amounts of components of the translation apparatus: ribosomes , tRNA , concomitant enzymes and protein factors. Moreover, unstable cell components, such as mRNA and nucleoside triphosphates , are absent, but there are reserves of stable ATP precursors - ADP and AMP [6] .
The complex system of endospore integuments makes it highly resistant to various adverse factors. Its protein coat blocks access to the cortex by enzymes that destroy peptidoglycan (for example, lysozyme ), and also protect spores from aggressive chemicals, such as chloroform . Membranes of sporangia and spores create a barrier of permeability around the core, preventing hydrophilic compounds weighing more than 300 Da from passing through it. In addition, the low water content in the cortex prevents the genomic DNA . Cations of divalent metals (primarily calcium ) are chelated by molecules , which are synthesized in sporangia and then delivered to the core, which provides additional protection for DNA. In the endospore cortex of some bacteria, there are special proteins that bind to DNA and protect it not only from depurinization, but also from the destruction of the saccharophosphate backbone. Finally, the germination of the endospore is accompanied by intensive repair of DNA damage accumulated during the resting period [7] .
Sporulation
The spore formation cycle takes much longer (about 7 hours at 37 ° C) than division with the formation of other specialized cells. In Bacillus subtilis, the sporulation cycle includes seven stages (0 – VII), which were proposed in the 1960s by the French scientist Antoinette Riter.
- Stage 0 . The original vegetative cell.
- Stage I. At present, this stage is not distinguished, since it is devoid of characteristic morphological characters, and mutants that are not able to pass it are unknown.
- Stage II is divided into three stages. Separation occurs at stage IIi: a spore septum forms in the mother cell and asymmetric binary division occurs, with it protoplasts of different sizes are formed. At stages IIii – IIiii, the larger protoplast begins to absorb the smaller one.
- Stage III . Absorption is completed, and a smaller protoplast (prespore) is formed in the cytoplasm of a larger cell (sporangia).
- Stage IV . A synthesis of the modified cell wall, the cortex, that surrounds the prespore occurs.
- Stage V. In the cytoplasm of sporangia, the protein integument of the future endospore is deposited.
- Stage VI . The stage is not morphologically expressed; during it, the endospores mature. It acquires increased resistance to environmental factors, enters into hyperanabiosis, becomes capable of germination.
- Stage VII . Sporangia undergoes programmed death and is partially lysed , which is why the mature endospore comes out [8] .
A unique spore septa is formed upon the symmetrical formation of two Z-rings on ¼ and ¾ of the mother cell. One of them is randomly selected with the participation of the SpoIIA protein, the second is destroyed. Further, in the region of the Z-ring, as in normal division , a circular invagination of the membrane occurs, which is subsequently filled with peptidoglycan . Then it is lysed, and as a result, the sister compartments are separated by only two membranes. The chromosome of the future endospore, according to recent data, penetrates into the smaller compartment after the closure of the septum. Its transfer is mediated by the DNA FtsK / SpoIIIE, which forms two through circular hexameric channels in the compartment membranes, and the C-terminal domains of the tubular proteins supply the process with ATP energy . The process of absorption of a smaller compartment by a large one generally corresponds to endocytosis (which is generally not characteristic of bacteria), its molecular mechanisms have been poorly studied [9] .
In the laboratory, bacteria begin to sporulate when an easily metabolized source of carbon , nitrogen, or phosphorus ends in the environment, but sporulation triggers in the bacteria’s natural habitat are unknown. A key event in the onset of sporulation is the phosphorylation of transcription factor Spo0A using protein kinases . Spo0A phosphorylation can be triggered in three ways:
- Through the kinase KinA, which records the redox and energy status of the cell.
- Through a decrease in the level of HDF and GTP in the cell, which indicates a lack of nutrients .
- Through proteins associated with a sense of quorum , that is, when a certain population density of bacteria is reached [10] .
Under conditions not conducive to sporulation, phosphorylated Spo0A is dephosphorylated by Spo0E phosphatase . Active phosphorylated Spo0A activates the transcription of sporulation regulators SpoIIA, SpoIIE and SpoIIG. Cells containing phosphorylated Spo0A secrete proteins that kill neighboring cells with inactive Spo0A in order to use their resources for sporulation. Also, the sigma factor σ H , which controls the spoIIID and spoIIIM genes , plays a role in triggering sporulation. Their products are involved in the formation of a spore septa. After the formation of the septum, the activity of Spo0A increases in sporangia, but decreases in prepo. Then, the sigma factor σ F is activated in the perspiration, and σ E in the sporangia. Also, in the prepore, σ G is synthesized, which is activated only upon completion of absorption, and in the sporangia at the final stages of spore formation, the sigma factor σ K is synthesized. Together, the sigma factors of prespore and sporangia activate the expression of genes necessary for the formation of endospores [11] .
Dormancy and Germination
The resting endospore is characterized by hyperanabiosis and hyperresistance. It does not exhibit metabolic activity, does not contain the most important metabolites , such as ATP and acetyl-CoA , the enzymes in it are inactive. In a state of hyperanabiosis, spores can remain viable over an enormous period of time. Thus, anthrax endospores in cattle burial grounds remain viable for 500 years, actinomycete spores - up to 7500 years [12] . There is evidence that spores of Bacillus sp. extracted from crystals of salt in New Mexico , remained viable for 250 million years. Endospores do not die under the action of high and low temperature , during drying, high hydrostatic pressure , under UV and γ radiation , under the influence of strong oxidizing agents , with increased acidity and other adverse conditions [13] . Spores of some bacteria even withstand boiling for an hour or more; therefore, solutions and instruments are sterilized in autoclaves with sterilization temperatures up to 121 ° C [12] .
Under favorable conditions, the endospore sprouts, that is, it leaves sporangia and turns into a normal vegetative cell. Germination inducers can be both physiological (some amino acids and sugars , purine nucleosides and other compounds or mixtures thereof), and non-physiological (mineral salts , exogenous calcium dipicolinate, lysozyme, cationic detergents , sublethal , pressure from 100 to 600 MPa ). Germination begins in seconds after exposure to the inductor and then does not depend on it. First, protons , potassium , sodium and zinc ions , dipicolinic acid with chelated Ca 2+ ions come out of the endospore, water enters. Then, with the help of special enzymes, the cortex peptidoglycan is lysed, rehydration continues, in which the core swells and the wall stretches. Finally, normal metabolism and biosynthesis of macromolecules resumes [14] .
Visualization
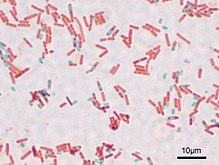
Endospores are clearly visible using both light and electron microscopes . Since endospores are impervious to many dyes , they are visualized as unpainted bodies against the background of the rest of the stained contents of the bacterial cell. However, there are methods for , by which spores become visible under a light microscope as blue bodies in a pink cytoplasm [12] .
Notes
- ↑ Pinevich, 2009 , p. 257.
- ↑ Pinevich, 2009 , p. 258-259.
- ↑ Pinevich, 2009 , p. 258.
- ↑ Pinevich, 2009 , p. 257-258.
- ↑ Netrusov, Kotova, 2012 , p. 67-68.
- ↑ Netrusov, Kotova, 2012 , p. 68-69.
- ↑ Pinevich, 2009 , p. 261-262.
- ↑ Pinevich, 2009 , p. 263.
- ↑ Pinevich, 2009 , p. 263-264.
- ↑ Pinevich, 2009 , p. 266-267.
- ↑ Pinevich, 2009 , p. 267-268.
- ↑ 1 2 3 Netrusov, Kotova, 2012 , p. 67.
- ↑ Pinevich, 2009 , p. 259-261.
- ↑ Pinevich, 2009 , p. 268-269.
Literature
- Pinevich A.V. Microbiology. Biology of prokaryotes: in 3 volumes - St. Petersburg. : Publishing House of St. Petersburg University, 2009. - T. III. - 457 p. - ISBN 978-5-288-04894-4 .
- Netrusov A.I., Kotova I. B. Microbiology. - 4th ed., Revised. and add. - M .: Publishing Center "Academy", 2012. - 384 p. - ISBN 978-5-7695-7979-0 .
- Modern Microbiology / Ed. J. Langelera, G. Drevs, G. Schlegel. - M .: Mir, 2005 .-- T. 1 .-- 654 p.